
![]()

(Photo by H. Tanaka)
この部門は時代に則した学際的共同研究を推進するために設立された新しい研究室です。色々な研究領域の人が推進者となって計画した研究テーマを選抜し、色々な解析手段を用いて共同プロジェクトとして6年間で遂行するものです。平成16年度からは生物多様性・保全生物学の研究を推進することになりました。これは遺伝学、行動学、生態学、生理学、形態学の視点と技術を用いて、特にアジア霊長類の生物多様性ならびに保全にかかわる研究を行うものです。
助手:香田啓貴(こうだひろき)(音声コミュニケーション)
非常勤研究員:早野あづさ(分子系統生物地理学)
推進代表者:平井啓久(遺伝子情報分野)、正高信男(認知学習分野)
推進者:景山節(人類進化モデル研究センター)、川本芳(集団遺伝分野)、松林清明(人類進化モデル研究センター)、室山泰之(ニホンザル野外観察施設)、高井正成(系統発生分野)、田中洋之(集団遺伝分野)、渡邊邦夫(ニホンザル野外観察施設)
協力研究者:杉浦秀樹(社会構造分野)
協力大学院生:D2 親川千紗子(認知学習分野)
研究内容の紹介
現在主に行っている研究は、テナガザル類の種分化・亜種分化に関する分子細胞遺伝学的、分子遺伝学的ならびに行動学的(音声)解析です。各課題を勢力的に遂行しながら、その成果を学際的研究として統合することを目指しています。


プロジェクト1:テナガザル類の系統生物地理学的研究(早野あづさ、田中洋之、Hery Wijayanto, Dyah Perwitasari-Farajallah, Alan Mootnick、平井啓久)
テナガザル類は、絶滅危惧種としてCTES(絶滅のおそれのある野生動植物の種の国際取引に関する条約:ワシントン条約)の附属書 I に記載されている。高木原生林を本来の生息地とする彼らは、森林の伐採によってその生息環境を急激に失いつつある。そのため、テナガザル類を保護する試みが色々な地域で始まっている。
生物の保全計画を有効なものにするためには、やみくもに保全対策を実施するのではなく、「進化的に生じた生物多様性の構成単位」あるいは「進化遺産のまとまり」のような具体的指標に基づいて計画する必要がある。そのためには、生態学的観察だけでなく、集団や個体の遺伝的な背景を正確に捉えることが求められる。すなわち、保全生物学には、形態学による種分類と生息データに加えて、種の多様性や歴史的展開を理解するうえで最も重要な情報となる遺伝学的データが欠かせない。
最近、我々はアジルテナガザル類に特異な染色体変異(第8・9染色体間全腕点座)を発見し、予備的解析からその変異がスマトラのアジルテナガザル亜種だけに存在するらしいという推測をたてた。もし、それが正しいとすると、アジルテナガザルの亜種分化を認識する標識のひとつとして重要である。

アジルテナガザルの染色体転座
そこで、我々はインドネシア・ボゴール農科大学霊長類研究センターと米国テナガザル保全センターを共同研究者として、「アジルテナガザルの亜種分化に関する総合調査」というプロジェクトを組織した。現在もっとも良く使われている分類体系では、アジルテナガザルは、スマトラ島のトバ湖以南とマレー半島の一部に生息する低地アジルテナガザル(Haylobates agilis unko)、高地アジルテナガザル(H. agilis agilis)、およびボルネオ島(インドネシア領カイリマンタン州)に生息するボルネオシロホオヒゲテナガザル(H. agilis albibarbis)の3亜種が認められている。ボルネオ島にはカプアス川とバリト川に挟まれた地域に生息するH. a. albibarbis以外に、同じ亜属に属する近縁種のミューラーテナガザル(H. muelleri)が生息する。
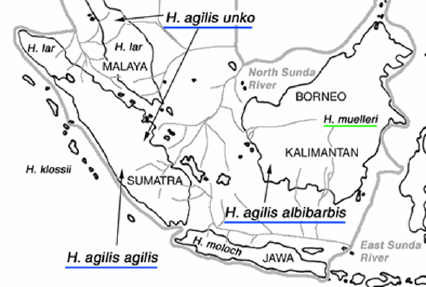
アジルテナガザルとミュラーテナガザルの分布
(1)早野あづさ
「テナガザル類の種分化・亜種分化に関する分子遺伝学的解析」
東南アジアの熱帯林に生息するテナガザル類は、類人猿の中では著しく多様性の高いグループです。このような生物グループの種分化および亜種分化の進化過程を明らかにすることは、多様性生物学の非常に興味深いテーマであるとともに、絶滅に瀕しているテナガザル類の多様性を保全するための基礎単位となる「進化的なまとまり」を認識するうえでも急務の課題といえます。
そのような研究を正確に行うためには、出自の明らかな個体について、多くの遺伝マーカーを用いて解析する必要があります。そこで、インドネシアとの共同研究プロジェクトを組織し、出自の明確な個体のDNA、染色体、形態、音声などの試料を収集しています。いくつかの核DNAの解析のひとつとして、マイクロサテライト遺伝子(約20座位)における繰り返し配列数の多型解析からテナガザル亜種間の遺伝的分化程度を評価しています。また、他の遺伝マーカー(染色体、Y染色体遺伝子、ミトコンドリアDNAなど)から得られたデータとの統合ならびにスンダ列島の地勢の歴史的変遷を考慮することによって、スマトラ・ボルネオ両島におけるテナガザルの地理的放散と分化の過程をより詳細に推定する予定です。
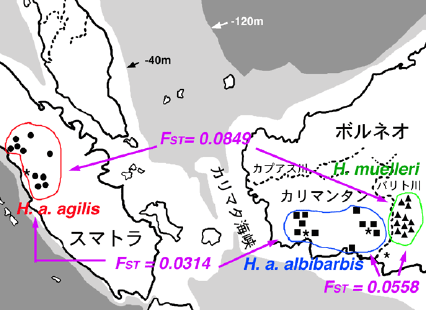
H. a. agilis, H. a. albibaribis, H. muelleri の遺伝距離(FST)
(2)Hery Wijayanto (インドネシア・ボゴール農科大学霊長類研究センター;ガジャマダ大学獣医学部)et al.
「テナガザル類、テナガザル属およびフクロテナガザル属、のC-ヘテロクロマチンとテロメア配列のパタン」: Patterns of C-heterochromatin and telomeric DNA in two representative groups of small apes, the genera Hylobates and Symphalangus
The course of chromosome evolution in small apes is still not clear, though painting analyses have opened the way for elucidating the puzzle. Even the C-banding pattern of the lar-group of gibbons (the genus Hylobates) is not clarified yet, although our previous studies suggested that lar-group gibbons have a unique C-banding pattern. We therefore made observations to establish C-banded karyotypes of the agile gibbons included in the lar group. The data were compared with those of siamangs (the genus Symphalangus), which carry distinctive C-bands, to determine the chromosomal patterns in each group. C-banded chromosomes of agile gibbons showed several terminal, interstitial, and paracentric bands, whose patterns are specific for each chromosome, whereas the C-bands of siamangs were located only at the terminal and centromeric regions in most chromosomes. Moreover, the C-bands of agile gibbons and siamangs were shown to be G+C-rich and A+T-rich DNA, respectively, by DAPI/C-band sequential staining. Additionally, PRINS labeling with a telomere primer revealed that agile gibbons have telomeric DNA only at chromosome ends where there is no C-band (nontelomeric heterochromatin), whereas the telomeric DNA of siamangs is located in the terminal C-banded regions (telomeric heterochromatin). Although the evolutionary mechanisms in small apes are still unknown, C-banding patterns and distribution of telomeric DNA sequences should provide valuable data to deduce the evolutionary pathways of small apes. (Chromosome Research, in press).
プロジェクト2:音声の研究(香田啓貴、親川千紗子、杉浦秀樹、正高信男)
東南アジアに生息するテナガザルの仲間は、ヒト、チンパンジー、ゴリラ、ボノボ、オランウータンと同じヒト上科というグループに属するサルの仲間です。同じグループの中でも、テナガザル類だけが多くの種に分かれています。研究者によって分類の仕方も色々で、14種に分類する研究もあります。種によって毛の色や体格など、行動も様々違います。
テナガザルの仲間に共通しているのは、オトナオスとオトナメスが夫婦となって一夫一妻のペアをつくることです。ペアにコドモ達が加わり3〜5頭ほどの家族を作り生活しています。
テナガザルたちのユニークな行動のひとつが「歌」と呼ばれる声です。サルの仲間はヒトと喉の構造が違っていたりするので、ヒトの言葉のような複雑な声を作ることができません。ところがテナガザルたちは、サルの仲間の中でもっとも複雑な声を作ることが知られています。ここではフクロテナガザル(シャーマン)、シロテテナガザル、ボウシテナガザル、アジルテナガザルの4種類のテナガザルを例としてあげましょう。
下の図は、4種類のテナガザルたちの声を図であらわしたものです。この図は「ソナグラム」と呼ばれ、縦に声の高さ、横に時間、色の濃さが音の強さを表しています。テレビなどで「声紋」と呼ばれているもので、ちょうど音楽の楽譜のようなものだと思ってください。彼らは、単純な「音素」と呼ばれる音を複雑に組み合わせて「歌」を歌います。テナガザルの仲間の多くは、この「歌」と呼ばれる声をオトナオスとオトナメスが鳴きあって「デュエット」と呼ばれる音声行動が見られます。このデュエットの役割については、いろいろな議論があり、まだまだ明らかにされていません。今のところ、夫婦の絆を強める役割があるといわれていますが、家族のなわばりを守る役割があるとも言われています。
歌やデュエットのやり方は、種によって様々です。フクロテナガザルはとても大きな声をだし、オスとメスがいつも一緒に鳴いています。しかし、ルールがあって図の矢印で示したような声はオトナオスしか鳴きません(「アーッ」と耳に聞こえる声です)。また喉袋を膨らまして低い音をだすこともフクロテナガザルの特徴です。シロテテナガザル、アジルテナガザル、ボウシテナガザルはメスが中心となってデュエットを歌います。特に、メスがもっとも盛り上がって鳴いている10秒間ほどの歌を「グレートコール」と呼びます(図1)。フクロテナガザルともっとも違う点は、メスがグレートコールを鳴いている間はオスは鳴かないことです。グレートコールをメスが鳴き終わってから、オスは「コーダー」と呼ばれる返事の歌を鳴きます。この鳴く順番の役割交代はこの3種の仲間の特徴といえるでしょう。さらに、図2、3、5のグレートコールを良く見てください。シロテテナガザル、アジルテナガザルはとても歌の特徴が似ていますが、ボウシテナガザルはとても違います。
同じテナガザルの仲間でも、メスとオスの歌の役割分担、歌の特徴など驚くほど違うのです。どのようにこの歌の違いが生まれたのか、現在研究が進められています。

******************************************************************************************
本ページに関するお問い合わせは(平井啓久)まで。